
Характерные для эволюции популяций «популяционные волны» или вспышки и деградации численности – генетик-эволюционист С.С. Четвериков [1, 2] назвал «волнами жизни» и это явление фундаментально для всех видов организмов.
К примеру, Э.Н. Фалалеев [3] выявил периодичность роста пихтовых насаждений по запасам древесины (Рисунок 1), что очень грубо, сглажено и косвенно отражает периодическую динамику численности пихты по неким когортам поколений.
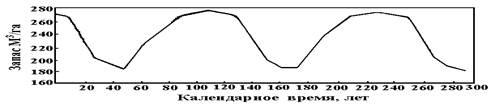
Рисунок 1. Периодичность запаса пихты на 1 га
В большинстве случаев, у организмов нельзя выделить чистые поколения, так как, к примеру, у человека новорожденный может родиться в любой момент времени, как и у микроорганизмов, размножение может произойти в любой момент времени, а у растений из-за разных сроков прорастания (стратификации) семян можно выделить только некие смешенные когорты поколений.
Для уточнения явления периодичности численности древесных пород по неким когортам поколений, в еловой популяции Рузского лесхоза Московской области (1991 г.) нами было заложено 8 больших пробных площадей в разных выделах и эти пробы были объединены в единый массив данных – 1810 деревьев ели (Рисунок 2). Так как, это периодическое явление присуще всей популяции ели (Рисунок 1) [1, 2, 3, 4, 5, 6].
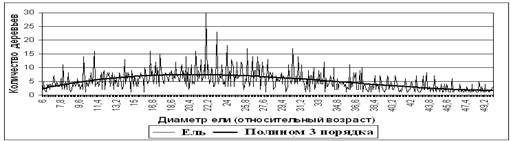
Рисунок 2. Численность деревьев ели по когортам поколений
На пробных площадях измерялась окружность дерева на высоте 1,3 м и преобразовывалась в точный диаметр. Среднегодовой прирост ели второго бонитета примерно равен 2,76*2 мм (0,552 см) диаметра [7], что позволяет рассчитать примерные возрастные периоды когорт поколений в годах, а для более точного определения календарного возраста деревьев его надо выяснять на вырубках по годичным кольцам.
Из исходного ряда данных (Рисунок 2) вычленялся тренд – полином 3 порядка, и к ряду всех ± остатков добавлялась положительная величина «А», равная самому большому отрицательному остатку, что бы ряд остатков стал положительным, так как спектральный статистический анализ [8] проводится только с положительными значениями данных. Все ряды остатков не сглаживались средне–скользящими, и по ним производился автокорреляционный и спектральный статистический анализ (Рисунок 3, 4).
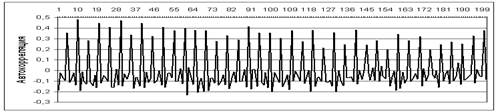
Рисунок 3. Автокорреляция остатков от тренда численности деревьев ели
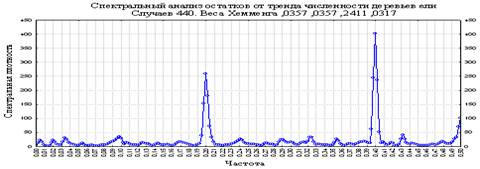
Рисунок 4. Спектральная плотность и частота остатков от тренда количества деревьев ели
На Рисунке 2 присутствуют два тренда – полином 3 порядка и тренд изменения «дисперсии» отклонений. Чтобы вычленить оба этих тренда, исходный ряд данных преобразовывался в проценты от тренда-функции. При этом, тенденция периодичности осталась та же, но достоверность результатов возросла (Рисунки 4 и 5, Таблица 1). А также проводился анализ периодичности средних признаков фено-форм древесных пород, различной реакцией на внешние физико-биологические факторы среды и т.п. Спектральный анализ проводился в абсолютных и относительных остатках от тренда. Во всех таблицах все достоверные значения выявленной периодичности заштрихованы.
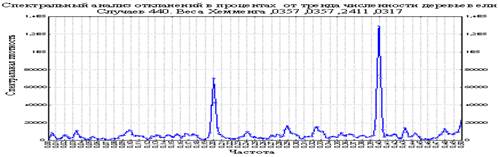
Рисунок 5. Спектральная плотность и частота количества ели в процентах от тренда
Таблица 1.
Результаты спектральных анализов по когортам поколений в популяции ели
|
Частота
|
Период диаметра (см) |
Период, лет
|
Отклонение в процентах от тренда |
Остатки от тренда |
||||||
|
Косинус коэф- фициент |
Синус коэф- фициент |
Периодо- грамма
|
Плотность
|
Косинус коэф- фициент |
Синус коэф- фициент |
Периодо- грамма |
Плотность |
|||
|
Периодичность количества деревьев ели по когортам поколений. |
||||||||||
|
0,005 |
220,0 |
≈399 |
7,301 |
-5,096 |
17441 |
8689 |
0,325 |
-0,307 |
44 |
23 |
|
0,034 |
29,3 |
≈53 |
-9,534 |
1,439 |
20453 |
10912 |
-0,481 |
0,114 |
54 |
31 |
|
0,100 |
10,0 |
≈18 |
2,868 |
7,839 |
15330 |
11088 |
0,202 |
0,387 |
42 |
30 |
|
0,200 |
5,0 |
≈9 |
25,718 |
3,105 |
147633 |
70620 |
1,460 |
0,187 |
476 |
260 |
|
0,325 |
3,1 |
≈6 |
-8,592 |
2,665 |
17806 |
15237 |
-0,312 |
0,329 |
45 |
32 |
|
0,400 |
2,5 |
≈5 |
35,922 |
-0,005 |
283890 |
129577 |
1,896 |
-0,075 |
793 |
402 |
|
Периодичность степени плодоношения ели по когортам поколений. |
||||||||||
|
0,009 |
110,0 |
≈200 |
-4,071 |
0,242 |
2745 |
2493 |
-0,109 |
0,005 |
2 |
2 |
|
0,021 |
47,1 |
≈86 |
-3,646 |
2,947 |
3626 |
2159 |
-0,080 |
0,069 |
2 |
1 |
|
0,030 |
33,0 |
≈60 |
1,483 |
4,586 |
3832 |
2659 |
0,032 |
0,109 |
2 |
2 |
|
0,079 |
12,7 |
≈23 |
0,340 |
4,990 |
4127 |
2390 |
0,001 |
0,110 |
2 |
1 |
|
0,127 |
7,9 |
≈14 |
4,618 |
-0,388 |
3543 |
2987 |
0,108 |
-0,013 |
2 |
2 |
|
0,188 |
5,3 |
≈10 |
-1,449 |
-4,165 |
3209 |
2170 |
-0,041 |
-0,085 |
1 |
1 |
|
0,209 |
4,8 |
≈9 |
-3,559 |
-1,496 |
2459 |
2160 |
-0,075 |
-0,036 |
1 |
1 |
|
Периодичность грубости коры по когортам поколений в популяции ели |
||||||||||
|
0,006 |
166,0 |
≈301 |
1,622 |
7,621 |
10078 |
6994 |
0,018 |
0,170 |
5 |
3 |
|
0,036 |
27,7 |
≈50 |
-4,419 |
5,435 |
8146 |
5952 |
-0,047 |
0,111 |
2 |
2 |
|
0,096 |
10,4 |
≈19 |
-5,326 |
-3,777 |
7077 |
4120 |
-0,091 |
-0,071 |
2 |
1 |
|
0,111 |
9,0 |
≈17 |
-1,865 |
-6,178 |
6912 |
6113 |
-0,051 |
-0,147 |
4 |
4 |
|
0,157 |
6,4 |
≈12 |
-6,648 |
-2,686 |
8534 |
4429 |
-0,144 |
-0,062 |
4 |
2 |
|
0,262 |
3,8 |
≈7 |
5,287 |
5,998 |
10613 |
6253 |
0,115 |
0,121 |
5 |
3 |
|
0,349 |
2,9 |
≈5 |
6,067 |
3,135 |
7742 |
4542 |
0,116 |
0,044 |
3 |
2 |
|
Периодичность относительного размера кроны деревьев ели по когортам поколений |
||||||||||
|
0,016 |
60,8 |
≈110 |
9,325 |
1,432 |
13528 |
7754 |
0,252 |
0,026 |
10 |
5 |
|
0,053 |
19,0 |
≈35 |
-5,740 |
2,228 |
5762 |
4326 |
-0,152 |
0,055 |
4 |
3 |
|
0,076 |
13,2 |
≈24 |
1,839 |
-4,234 |
3238 |
2213 |
0,041 |
-0,117 |
2 |
1 |
|
0,092 |
10,9 |
≈20 |
-0,280 |
-7,737 |
9111 |
5550 |
-0,026 |
-0,211 |
7 |
4 |
|
0,118 |
8,4 |
≈16 |
-5,099 |
-2,434 |
4852 |
3319 |
-0,141 |
-0,067 |
4 |
2 |
|
0,128 |
7,8 |
≈14 |
1,536 |
7,099 |
8019 |
4325 |
0,037 |
0,180 |
5 |
3 |
|
0,214 |
4,7 |
≈9 |
4,909 |
-2,676 |
4751 |
3279 |
0,116 |
-0,077 |
3 |
2 |
|
0,411 |
2,4 |
≈5 |
1,868 |
-4,812 |
4050 |
2082 |
0,037 |
-0,105 |
2 |
1 |
Первый столбец Таблицы 1 отражает относительную частоту ряда данных.
Второй столбик отражает период, выраженный в сантиметрах диаметра дерева.
Третий столбец отражает примерный календарный период, рассчитанный на основании второго столбика и таблиц хода роста по породе [7]. Ход роста по диаметрам древесных пород в естественных насаждениях подчиняется логистической регрессии и средняя часть этой функции практически прямолинейная, а искривления в начале и конце этой кривой, хотя и вносят систематические ошибки, но они явно незначительны, иначе бы мы, не смогли-бы выявить достаточно детерминированные цикличности.
Косинус и Синус коэффициенты отражают – есть ли склоны слева и справа у выявленного среднего цикла, так как регулярная периодичность может быть и в виде регулярных всплесков без каких-либо склонов.
Периодограмма – оценка спектральной плотности мощности – функция, задающая распределение мощности отклика признака по частотам и имеет размерность мощности делённой на частоту.
Спектральная плотность – это функция, которая показывает распределение мощности признака по частотам.
Плодоношение ели выражено в баллах: 1 балл – нет плодоношения; 2 – слабое плодоношение; 3 – среднее плодоношение; 4 – сильное плодоношение.
Характеристика коры ели выражена также в баллах: 1 балл – кора гладкая; 2 – полугладкая; 3 – полугрубая; 4 – грубая.
Относительный размер кроны – это отношение высоты дерева к высоте до первой нижней живой ветви и косвенно отражает объемы биохимических процессов. К примеру, на простом графике, встречаемость скорости работы печени (соответственно и объемов биохимических процессов организма) по популяции человека имеет двуглавую зависимость [8]. Это может объяснено тем, что в единую выборку необъективно объединены две разные генеральные распределения совокупности этого признака, характеризующие разные по этим свойствам (четные и нечетные) когорты поколений, что обязательно следует учитывать, если выборка объединяет разные когорты поколений популяции.
Средневзвешенная категория состояния кроны популяции ели в 1991 г. равна 1,34 и некий очень малый процент усыхания ветвей, встречался только на угнетенном подросте 4 и 5 класса Крафта и поэтому спектральный анализ по этому признаку не проводился.
По всем Тульским засекам (1993 г.) заложены пункты учёта мониторинга. Наблюдения объединены в единые массивы данных – 690 деревьев дуба и 1002 липы. Измерялась окружность дерева на высоте 1,3 м. Средний прирост дуба второго бонитета равен 2,8*2 мм [7] или 1,76 см окружности. Таблицы хода роста по липе в литературных источниках отсутствуют и, поэтому, периоды приводятся только по окружности деревьев (Таблица 2).
У всех древесных пород выявлен индивидуально-неповторимый комплекс гармоник цикличности по численности поколений и у всех проанализированных полигенных признаков (Таблица 1, 2) – сравнивай второй столбец между всеми полигенными признаками по каждой породе, а по 3 столбику соответствующие признаки между породами.
Таблица 2.
Результаты спектральных анализов популяции дуба и липы
|
Частота |
Окружность (см) |
Период лет |
Отклонение в процентах от тренда |
Остатки от тренда |
||||||
|
Косинус коэф- фициент |
Синус Коэф- фициент |
Периодо- грамма |
Плотность |
Косинус коэф- фициент |
Синус коэф- фициент |
Периодо- грамма |
Плотность |
|||
|
Периодичность количества деревьев дуба по когортам поколений |
||||||||||
|
0,018 |
56,0 |
≈32 |
1,647 |
8,908 |
6893 |
3608 |
0,136 |
0,382 |
14 |
8 |
|
0,095 |
10,5 |
≈6 |
-8,831 |
-9,756 |
14545 |
9632 |
-0,220 |
-0,574 |
32 |
21 |
|
0,149 |
6,7 |
≈4 |
-6,195 |
-7,688 |
8189 |
5012 |
-0,128 |
-0,430 |
17 |
11 |
|
0,250 |
4,0 |
≈2 |
7,740 |
-8,626 |
11283 |
7369 |
0,240 |
-0,308 |
13 |
9 |
|
0,440 |
2,3 |
≈1 |
10,733 |
-3,214 |
10543 |
5598 |
0,554 |
0,007 |
26 |
15 |
|
Периодичность процента усыхания кроны деревьев дуба по когортам поколений |
||||||||||
|
0,039 |
25,7 |
≈15 |
-6,080 |
-8,632 |
8584 |
7129 |
-0,453 |
-1,066 |
103 |
84 |
|
0,065 |
15,4 |
≈9 |
-3,223 |
13,382 |
14590 |
8825 |
-0,318 |
1,141 |
108 |
63 |
|
0,110 |
9,1 |
≈5 |
-8,339 |
-14,297 |
21093 |
15445 |
-0,409 |
-1,195 |
123 |
87 |
|
0,143 |
7,0 |
≈4 |
-0,351 |
-15,032 |
17408 |
10961 |
-0,149 |
-1,225 |
117 |
73 |
|
0,273 |
3,7 |
≈2 |
9,786 |
10,812 |
16375 |
13954 |
0,864 |
1,026 |
139 |
137 |
|
0,435 |
2,3 |
≈1 |
-18,310 |
-3,852 |
26957 |
15404 |
-1,976 |
-0,331 |
309 |
193 |
|
Периодичность процента объедания кроны деревьев дуба по когортам поколений |
||||||||||
|
0,019 |
52,0 |
≈30 |
9,070 |
-7,441 |
10736 |
10564 |
0,818 |
-0,852 |
109 |
123 |
|
0,045 |
22,3 |
≈13 |
-8,954 |
14,792 |
23319 |
12690 |
-1,017 |
1,633 |
289 |
161 |
|
0,135 |
7,4 |
≈4 |
-15,174 |
-15,066 |
35664 |
24974 |
-1,701 |
-1,301 |
358 |
252 |
|
0,173 |
5,8 |
≈3 |
15,340 |
6,910 |
22080 |
12853 |
1,474 |
0,988 |
246 |
132 |
|
0,353 |
2,8 |
≈2 |
-0,591 |
-14,425 |
16259 |
10792 |
-0,077 |
-1,760 |
242 |
157 |
|
0,500 |
2,0 |
≈1 |
19,274 |
0,000 |
28975 |
17665 |
1,861 |
0,000 |
270 |
156 |
|
Периодичность количества деревьев липы по когортам поколений |
||||||||||
|
0,025 |
40,5 |
|
8,820 |
4,154 |
7698 |
6619 |
0,336 |
-0,013 |
9 |
15 |
|
0,056 |
18,0 |
|
0,982 |
-9,831 |
7907 |
5193 |
0,029 |
-0,342 |
10 |
7 |
|
0,093 |
10,8 |
|
8,741 |
4,206 |
7622 |
4853 |
0,386 |
0,318 |
20 |
12 |
|
0,216 |
4,6 |
|
8,543 |
-5,376 |
8253 |
7098 |
0,141 |
-0,350 |
12 |
17 |
|
0,321 |
3,1 |
|
3,282 |
-6,828 |
4648 |
6387 |
0,053 |
-0,422 |
15 |
22 |
|
0,444 |
2,3 |
|
7,395 |
7,558 |
9057 |
7012 |
0,379 |
0,343 |
21 |
17 |
|
Периодичность процента тиростромоза деревьев липы |
||||||||||
|
0,013 |
79,0 |
|
52,828 |
-2,485 |
220958 |
209187 |
1,289 |
1,634 |
342 |
202 |
|
0,051 |
19,8 |
|
41,835 |
-0,587 |
138293 |
118788 |
1,316 |
-0,438 |
152 |
128 |
|
0,089 |
11,3 |
|
41,318 |
-9,505 |
142006 |
116060 |
0,921 |
-1,441 |
231 |
156 |
|
0,133 |
7,5 |
|
37,188 |
-5,318 |
111486 |
85508 |
0,812 |
-0,872 |
112 |
67 |
|
0,203 |
4,9 |
|
38,363 |
-4,268 |
117707 |
86021 |
0,883 |
-1,101 |
157 |
121 |
|
0,234 |
4,3 |
|
35,911 |
9,630 |
109203 |
93920 |
0,332 |
0,562 |
34 |
43 |
|
0,342 |
2,9 |
|
37,744 |
5,632 |
115053 |
94934 |
0,603 |
0,210 |
32 |
28 |
|
Периодичность процента объедания кроны деревьев липы |
||||||||||
|
0,038 |
26,3 |
|
-8,144 |
-2,447 |
5712 |
3792 |
-1,012 |
-0,453 |
97 |
68 |
|
0,070 |
14,4 |
|
-18,616 |
2,294 |
27792 |
17924 |
-2,496 |
0,194 |
495 |
344 |
|
0,114 |
8,8 |
|
0,443 |
-13,410 |
14223 |
11487 |
-0,064 |
-1,404 |
156 |
135 |
|
0,133 |
7,5 |
|
-9,266 |
-6,494 |
10115 |
8527 |
-1,402 |
-0,973 |
230 |
170 |
|
0,165 |
6,1 |
|
0,861 |
11,030 |
9671 |
9989 |
0,136 |
1,317 |
139 |
130 |
|
0,209 |
4,8 |
|
1,171 |
17,115 |
23248 |
13738 |
0,255 |
2,291 |
420 |
272 |
|
0,310 |
3,2 |
|
-9,001 |
4,860 |
8267 |
4827 |
-1,083 |
0,705 |
132 |
82 |
В прошлом автокорреляционным анализом проанализировано 57 признаков 5 древесных пород [4] и выявлено, что каждый полигенных признак имеет свою сложную периодичность с множеством гармоник цикличности по неким когортам поколений.
Моногенные признаки организма контролируются одной парой аллельных генов, и они представлены как альтернативные признаки «есть – нет или такие – другие». Г.И. Мендель [10] нашёл математическую зависимость наследования моногенных признаков, даже не подозревая о существовании генов. Считается, что всех моногенных признаков у всех организмов не более 1%, а все остальные признаки являются полигенными.
Полигенные признаки организма контролируются многими генами, находящиеся в разных участках хромосомы, а иногда даже и в разных парах хромосом. Все полигенные признаки – метрические и их можно как-либо померить.
К примеру, более темные или светлые голубые глаза, или мера интеллекта, иммунитета, скорость и объем биохимических процессов, метрические различия фенотипической изменчивости или той или иной сексуальности выраженной вторичными половыми признаками – тем или иным гормональным балансом и т.п.
Периодичность численности любой популяции – это следствие периодичности неких сексуальных полигенных признаков и резонанса их гармоник цикличности. К примеру, по древесным породам, периодичная численность может зависеть от периодического качества и степени плодоношения, периодичности всхожести, приживаемости, смертности и т.п. полигенных признаков и это выражается в периодической изменчивости полового индекса. На однодомных растениях по когортам поколений также могут в большей или меньшей мере встречаться женские или мужские цветки, отражая их, тот или иной гормональный баланс. Вероятно, именно некая периодическая смена той или иной сексуальности «Сексуальные революции» и вызывают периодические вспышки и деградации численности в популяциях видов, как растений, насекомых, так и животных.
Демограф З. Коллер [11] утверждал, что «следующие друг за другом поколения новорожденных не обладают, и не могут обладать одинаковой жизненной силой: есть более жизнеспособные когорты поколений и менее жизнеспособные». А всем широко известно, что женские организмы или более женственные по вторичным половым признакам – более жизнеспособны, чем более мужские.
По автокорреляционному анализу (Рисунок 3) в соответствии с Таблицей 1 выявлен 10 см (≈18 лет) экзогенные цикл количества деревьев. Так как 5 см (≈9 лет) циклы модулированы и эта цикличность имеет череду увеличения и уменьшения амплитуды именно 5 см циклов через каждые 10 см (Рисунок 3), что можно объяснить только влиянием периодичности некого внешнего (экзогенного) фактора.
Примером тому может служить активность солнечных пятен [12], где есть «эндогенный» 11-летний цикл – со своей внутренней причиной Солнца. «Экзогенный» же цикл в 22 года имеет другую причину, связанную со сменой магнитных полюсов на Солнце, и это как «внешний» фактор модулирует, увеличивая и уменьшая амплитуды именно 11 летних циклов. Вековой экзогенный цикл солнечной активности также проявляется в модуляциях, уменьшая и увеличивая амплитуды именно 11-летних циклов. Выделяют и большие экзогенные цикличности Солнца и, вместе с вековым циклом, они, вероятно, также определяются какими-то экзогенными – космологическими факторами, подобные периодическому параду планет, галактик и т.п.
Следовательно, по форме цикличности можно различить эндогенные и экзогенные периоды, но спектральный статистический анализ этого не различает и надо разработать соответствующий статистический алгоритм. Эндогенные и экзогенные периодичности можно будет вычленять и раздельно анализировать их корреляционные и функциональные связи с эндогенными причинами и внешними (экзогенными) факторами.
Форму циклов, их начало, и конец можно различить и на простых графиках, но большинство точек на Рисунке 2 представлены 5ю деревьями или меньше. Соответственно, каждая частная средняя всех полигенных признаков по поколениям не является достаточно точной, но для выявления достоверной периодичной зависимости этих данных оказалось достаточно. Следует увеличить выборку в десятки раз, чтобы каждая точка графика (Рисунок 2) была наиболее достоверной.
При лесопатологическом мониторинге требуется расслоить популяцию древесных пород по окружностям (диаметрам) деревьев на особо чувствительные (сигнальные) когорты поколений к различным патологиям и менее чувствительные и рассматривать их статистически раздельно. При этом необязательно знать реальные календарные даты когорт поколений и эндогенные причины этих периодичностей.
В каждый момент времени в конкретной популяции должна быть уникально-согласованная взаимосвязь всех гармоник цикличности всех полигенных признаков в процентных пропорциях. Но так как когорты поколений нами определяются через окружность (диаметр) деревьев, а известно, что средний прирост пород зависит от условий местопроизрастания. Эту систематическую ошибку можно исключить между двумя большими пробными площадями и после этого объединить их данные. Надо заложить большую эталонную пробную площадь по конкретной древесной породе. Взять любой, наиболее вариабельный полигенный признак и рассчитать его цикличности на простом графике и совместить эту эталонную периодичность с соответствующими результатами другой большой пробы. При этом надо создать простой программный алгоритм рекуррентных итераций (сдвига лага периода влево – право и с расширением – сужением лага периода) и добиться того, чтобы эти периодические тенденции совпали, и критерием их максимального совпадения будет являться максимально возможный коэффициент прямой корреляции. При этом, до максимума возрастет прямой коэффициент корреляции и по любым другим периодичностям всех полигенных признаков данной популяции.
Выявленная индивидуально-сложная периодическая зависимость наследования всех полигенных признаков по когортам поколений популяций, позволит спрогнозировать все полигенные явления биологической эволюции. Эти периодические явления позволят изучать периодичную активность ответственных за это генов и их комплексов.
По всей вероятности, длина эндогенных периодов цикличности как-либо зависят от сложности ответственных за это генетических комплексов. Которые могут быть более или менее активными и подчиняться «часовым механизмам» ДНК [12]. Этим объясняется периодичность вспышек численности организмов, синхронность модных мутаций в изолированных популяциях [14] и не детерминация результатов от поколения к поколению при поиске «вещества памяти» у плоских червей – Планарий [15].
К примеру, С.Ю. Маслов [16] выявил, что доминирование симметрично-функциональных областей мозга периодично в популяциях человека, и человечество периодически интеллектуально различно (полигенный признак) по поколениям, а следовательно, тоже самое должно быть и Планарий, но с другими периодами. С.Ю. Маслов выявил череду циклов периодичности среднего интеллекта человечества близкое к 50 годам, где в начале цикла ведущую роль играет интуиция гениальных научных догадок, эвристических рассуждений и преобладает абстракционизм в архитектуре, литературе, искусстве и т.п.
Н.Д. Кондратьев [17] выявил экономические циклы в 52±4 года, что согласуется с циклами периодичности интеллекта человечества. В начале экономического цикла также появляются новые технологии, изобретения и открытия, что косвенно подтверждает выводы С.Ю. Маслова. По сути, Л.Н. Гумилев [18] писал об этом же периодическом явлении интеллектуального расцвета и заката этносов, но о гармонике цикличности с большим периодом. Этим же объясняется смена реалистичных наскальных рисунков периода палеолита на схематично абстрактные рисунки в неолите – еще большая гармоника цикличности интеллекта человечества. Из чего, следует, что полигенная периодичность интеллекта в эволюции организмов и их следствия детерминировано, различаются по неким когортам поколений. Это может быть также связано с некой сменой средней сексуальности по вторичным половым признакам, так как известно, что женщины или мужчины в некоторой степени интеллектуально различаются.
Наиболее полный и точный статистический анализ этих и им подобных сложных периодических явлений любых полигенных признаков и патологий можно провести по популяции человека, относительно быстро собрав наиболее полную информацию в медицинских картотеках, где есть даты рождений, заболеваний и патологий.
С лесохозяйственной точки зрения, Лес или насаждение – это совокупность тех или иных сырьевых, пищевых и биологических ресурсов. Которые, как правило, измеряются в интегральных показателях – суммы площадей, в процентах или в реальных кубометрах ослабленной, поврежденной и усохшей древесины, тоннах и т.п.. При этом, все эти интегральные показатели вычисляются опосредовано и не являются математическим ожиданием и поэтому на их основании можно попытаться сделать только очень грубые, интуитивно-краткосрочные прогнозы в лесопатологии.
С биологической точки зрения, Лес – это совокупность различных популяций микро и макро организмов, существующих в тех или иных условиях окружающей среды. Последнее отчасти отражают лесоустроительные таксационные описания насаждений.
При наземном лесопатологическом мониторинге учет полигенных признаков средне – фонового состояния популяции, вычисленный через число особей, и может быть определен, практически с любой требуемой точностью. Эти средние показатели, а так же доли и проценты, вычисленные от числа организмов, являются математическим ожиданием и на этом основании можно сделать детерминированные краткосрочные, среднесрочные и долгосрочные всевозможные прогнозы во времени и пространстве на основании множественных регрессионных моделей, матричного и комбинированного методов прогноза с привлечением Геоинформационной системы (ГИС).
В ФБУ «Рослесозащита» пока только частично организован санитарный мониторинг, оперирующий интегрально-валовыми показателями. С ежегодной частичной оценкой и учетом санитарного состояния на выборочной части лесных территорий, и санитарного состояния уже возникших очагов вредных организмов.
Для подобных краткосрочных, среднесрочных и долгосрочных прогнозов патогенных организмов, живущих 1-2 года надо следить за чередой их поколений, проводя ежегодный средне – фоновый мониторинг полигенных признаков (половой индекс, индекс веса, размер, доля встречаемости, плодовитость и т.п.) по каждому году наблюдений в их резервациях.
Для краткосрочных, среднесрочных и долгосрочных прогнозов надо создавать соответствующие временные ряды средних полигенных признаков когорт поколений популяций древесных пород, поколений популяций их патогенов и хвое-листо грызущих вредителей. Параллельно создавая временные ряды санитарного состояния ослабленных, усохших и усыхающих насаждений, возникших очагов вредителей и потом искать функциональные и корреляционные зависимости между периодическим средне – фоновым лесопатологическим состоянием популяций и санитарным состоянием насаждений. Требуемую для этого информацию можно найти в библиотеках – научных статьях, диссертациях, в данных из экспедиционных отчетов и мониторинга, и эту информацию требуется оцифровать в базы данных и создать временные ряды санитарного состояния лесов и лесопатологического фонового состояния популяций хозяина и его патологий.
Так как закономерно – периодически по когортам поколений рождаются организмы с конкретным сочетанием разных полигенных признаков. Это позволит спрогнозировать по когортам поколений массовое появление различных фено-форм с тем или иным сочетанием фено – физиологических свойств и их реакцией на, те или иные физико-биологические факторы внешней среды. Выявленные селективно-прогнозируемые когорты поколений с особо ценными сочетаниями фено – физиологических свойств растений можно потом клонировать вегетативно в сельском и лесном хозяйстве или в некоторые годы имеет смыслы уделять особое внимание сбору семян, а в некоторые нет.
По когортам поколений организмов закономерно – периодически «создаются» различные сочетания всех их полегенных признаков. К примеру, у вьюрковых птиц с определенным периодом периодичности должен создаваться некий размер, а с другими периодами форма клюва, размер, толщина костей – головы, шеи, лап, крыльев и т.п., и тех или других комплексно сформированных фено-форм закономерно – периодически во множестве рождающихся в конкретной когорте поколения. Это же периодическое явление может вызвать закономерную половую изоляцию по тем же периодично-эндогенным причинам и этим закономерно спровоцировать появление новых видов и родов, что всё позволяет создать естественную периодическую систематику всех организмов [4, 5, 6].
Экзогенный же естественный отбор Ч.Р. Дарвина [19, 20, 21] только лимитирует и приспособляет эту закономерную эндогенно-периодическую эволюцию [4, 5, 6] к той или другой внешней среде, в той или иной мере «разрешая или запрещая» существование этих, закономерно – периодически сформировавшихся фено-форм или видов и родов. Естественный отбор внешней среды может отобрать только то, что уже эндогенно и закономерно существует в популяции, к примеру, в популяции березовой пяденицы всегда есть и светлые и темные особи и соответственно естественный отбор может отбирать только то, что по периодическим закономерно-эндогенным причинам уже эволюционно во множестве появилось. При этом, у изолянтов с изолированных островов имеется своя – изолированная эндогенно-периодическая родословная связь наследования поколений и потому они в итоге отличны от других изолянтов с других островов и континентов.
Библиографическая ссылка
Сафронов В.Н. ЭНДОГЕННАЯ И ЭКЗОГЕННАЯ ПЕРИОДИЧНОСТЬ ПОЛИГЕННЫХ ПРИЗНАКОВ ПО КОГОРТАМ ПОКОЛЕНИЙ ПОПУЛЯЦИЙ // Научное обозрение. Фундаментальные и прикладные исследования. 2018. № 1. ;URL: https://scientificreview.ru/ru/article/view?id=13 (дата обращения: 21.07.2026).
DOI: https://doi.org/10.17513/srfar.13